Physiopathologie de la douleur chronique par Bernard CALVINO
Bernard CALVINO
Professeur Honoraire de Neurophysiologie
On le sait tous il n’y a pas une douleur, mais des douleurs selon la nature du mécanisme, douleurs aiguës mais aussi douleurs chroniques lorsqu’elles se prolongent au delà de 3 mois.
La douleur aiguë est une expérience sensorielle et émotionnelle résultant des stimulations dites « nociceptives » c’est à dire de haute intensité, qui déclenchent une cascade d’évènements physiologiques conduisant à l’intégration des informations codant les différents aspects de la douleur.
Le prolongement dans le temps de la douleur aiguë conduit au développement d’une douleur chronique.
La douleur perd sa signification de signal d’alarme pour évoluer vers un syndrome chronique.
Les douleurs chroniques peuvent être en rapport avec plusieurs causes :
– Douleurs par excès de nociception (inflammation, sensibilisation des nocicepteurs, …)
– Douleurs neuropathiques (neuropathies périphériques consécutives à une lésion de nerfs sensoriels périphériques, ou neuropathies centrales, lésions de structures relais des voies de la douleur dans le système nerveux central.
– Douleurs mixtes, douleurs par excès de nociception et douleurs neuropathiques, par exemple dans le cas des douleurs cancéreuses.
– Douleurs nociplastiques (IASP (2018), autrefois répertoriées comme douleurs idiopathiques ou fonctionnelles, sans lésion ni cause objective qui résultent de la plasticité du système nerveux central susceptible de modifier les systèmes de contrôle de la douleur et d’engendrer ainsi des douleurs sans cause apparente (par exemple la fibromyalgie).
La douleur peut être modulée en fonction de la situation psychologique du sujet, mais aussi en fonction de son environnement (affectif, socio-culturel, ethnologique, religieux, …). Cette modulation résulte de la mise en jeu de contrôles inhibiteurs exercés par des structures spinales et supra-spinales (corticales ou sous-corticales) par l’intermédiaire des voies descendantes.
D’un point de vue physiologique, la cascade d’évènements conduisant à la genèse et à l’intégration des informations douloureuses, s’inscrit dans un chapitre de la neurophysiologie sensorielle, la somesthésie (qui regroupe la physiologie sensorielle de la sensibilité tactile cutanée mais aussi des muscles, des tendons, des articulations et des viscères) : elle répond au même schéma fonctionnel que toute les autres fonctions sensorielles (audition, vision, …).
La douleur nociceptive aiguë résulte de la mise en jeu d’une triade :
Lésion-inflammation-douleur.
Les informations nociceptives à l’origine de la douleur aiguë sont générées à la périphérie par la lésion qui va être à l’origine d’une inflammation : de nombreuses molécules, constituant la « soupe inflammatoire », sont synthétisées et libérées par les cellules lésées des tissus périphériques, les terminaisons nerveuses et les cellules immunocompétentes activées qui migrent dans le foyer inflammatoire et sont à l’origine de la composante inflammatoire de la douleur.
Ces molécules sont susceptibles d’activer et/ou de sensibiliser les nocicepteurs périphériques, du fait que leurs récepteurs sont exprimés par les terminaisons nerveuses des fibres périphériques nociceptives de petit diamètre des nerfs sensoriels (dans lesquels on trouve aussi les fibres non nociceptives de la sensibilité tactile légère de gros diamètre).
Ces fibres sont les axones des neurones sensoriels primaires dont le corps cellulaire est localisé dans le ganglion de la racine dorsale (GRD)
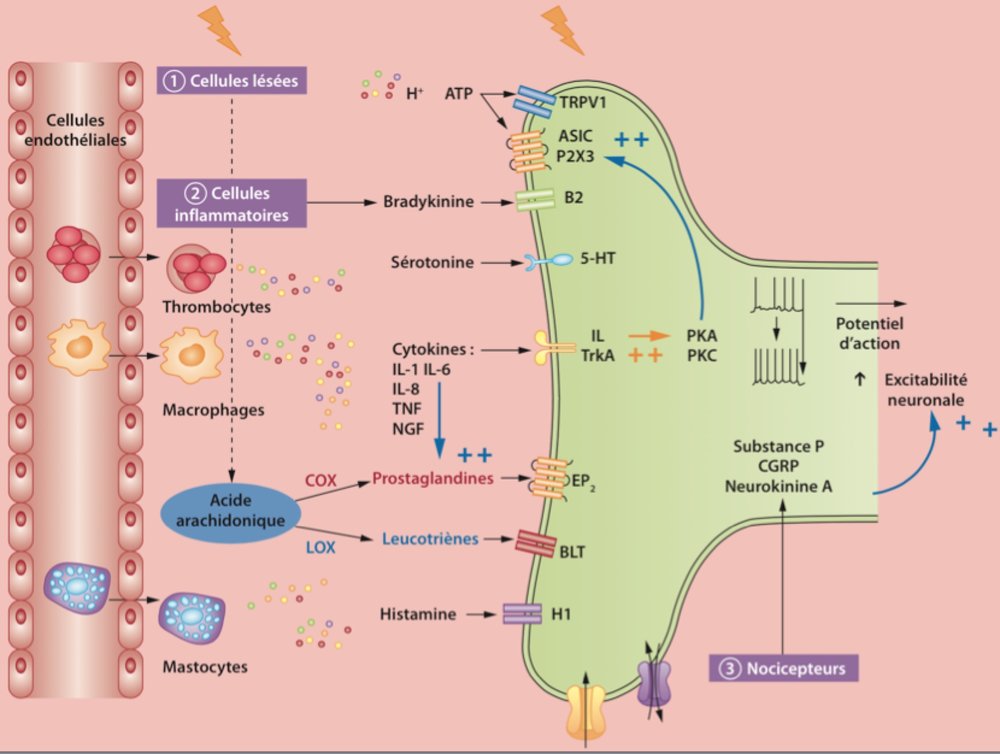
D’après Le Point Vétérinaire Juin 2015 n°356
Elles génèrent et conduisent l’information sensorielle jusqu’à la corne dorsale de la moelle épinière (CDME).
Ainsi, une stimulation nociceptive – stimulation de forte intensité générant une lésion de l’organisme – met ainsi en jeu les nocicepteurs périphériques.
Les nocicepteurs ne répondent qu’à des stimulations nociceptives (mécanonocicepteurs; thermonocicepteurs au chaud et au froid; chémonocicepteurs qui répondent aux molécules algogènes de la soupe inflammatoire; nocicepteurs polymodaux, qui répondent à plusieurs modalités de stimulations).
Dans la CDME, les axones des neurones de deuxième ordre (post-synaptiques), qui reçoivent les terminaisons centrales des neurones périphériques (neurones de premier ordre), les neurones nociceptifs post-synaptiques, sont à l’origine des voies médullaires appelées ascendantes parce qu’elles conduisent le message nerveux douloureux aux différents noyaux relais dans le cerveau, où sont intégrées ces informations douloureuses.
Trois classes de neurones nociceptifs post-synaptiques dans la CDME recevant des afférences des terminaisons des fibres sensorielles ont été identifiées :
1. Les neurones nociceptifs spécifiques ne répondent qu’à des stimulations périphériques nociceptives, ils reçoivent des afférences des fibres Aδ et C
2. Les neurones nociceptifs non spécifiques répondent à des stimulations périphériques de faible et de forte intensité, ils reçoivent des afférences de fibres sensorielles non nociceptives (fibres Aβ) et nociceptives (fibres Aδ, C)
3. Les neurones non nociceptifs spécifiques ne répondent qu’à des stimulations périphériques de faible intensité et ils reçoivent des afférences de fibres sensorielles non nociceptives (fibres Aβ), ils n’interviennent pas dans l’intégration de l’information nociceptive.
Les axones des neurones nociceptifs de la CDME constituent les faisceaux médullaires ascendants (dont les voies spinomésencéphalique (VSM) et spinoréticulée (VSR), qui projettent leur information à différents niveaux supraspinaux.
On distingue quatre sites supraspinaux de projection :
1. Les noyaux du thalamus ventro-postéro-latéral, (VPL) noyaux spécifiques de la sensibilité tactile et de la nociception, à l’origine de la composante sensori-discriminative de la douleur
2. Des sites de projection bulbaires (noyau gigantocellulaire) et mésencéphaliques (substance grise périaqueducale et noyau cunéiforme), structures relais pour l’information nociceptive véhiculée par le faisceau spino-réticulo-thalamique jusqu’au thalamus médian non spécifique
3. L’hypothalamus intervient dans le contrôle des réactions végétatives de la douleur, mais aussi dans la libération d’hormones intervenant dans le contrôle du stress
4. Le complexe amygdalien, structure du système limbique, intervient dans les réactions affectives et émotionnelles de la douleur.
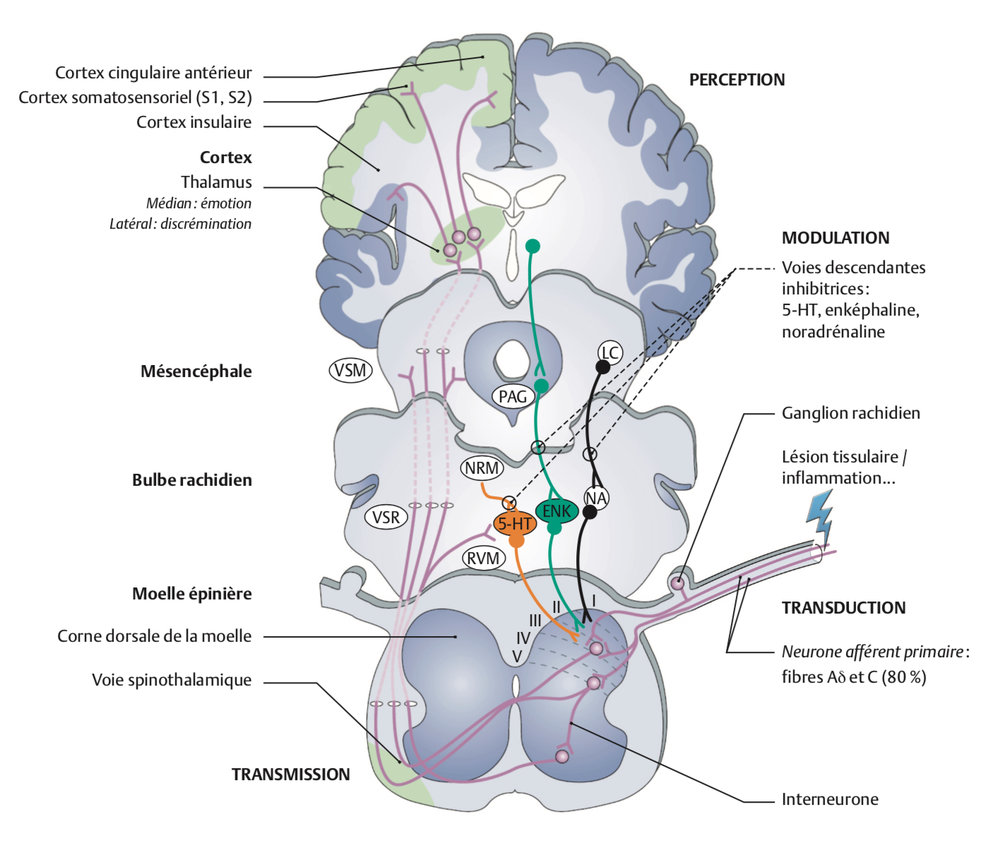
D’après Beaulieu Pierre modifié Fields: La douleur Guide pharmacologique et thérapeutique. Les presses de l’université de Montréal 2013
Les sites de projection corticaux sont multiples et les interactions entre ces sites sont nombreuses.
1. Les neurones du thalamus VPL projettent leurs axones vers les aires des cortex somesthésiques primaire S1 et secondaire S2 (cortex spécifiques de la somesthésie et de la nociception): les caractéristiques du message nociceptif y sont décodées permettant la genèse de la perception de la sensation douloureuse (qualité, localisation, intensité, durée).
2. Différents noyaux de l’encéphale impliqués dans la douleur projettent leurs axones vers les aires corticales pré-frontales, le cortex insulaire et le cortex cingulaire antérieur, impliqués dans les réactions émotionnelles plus élaborées à la douleur et qui jouent un rôle fondamental dans la composante émotionnelle et cognitive de la douleur, l’ensemble constituant « la matrice de la douleur ».
La complexité de la compréhension des bases neurophysiologiques de la douleur vient en grande partie de la multiplicité des voies ascendantes localisées dans la moelle épinière, des nombreuses structures de projection du tronc cérébral et de l’encéphale qui contribuent toutes à la genèse de la douleur et à sa modulation qui résulte de la mise en jeu de contrôles exercés par des structures spinales et supra-spinales.
On distingue quatre principales catégories de systèmes physiologiques de contrôle de la douleur.
Les contrôles segmentaires spinaux de la CDME
Les contrôles segmentaires spinaux émergent de la CDME qui est un lieu d’intégration et pas seulement un relais de transmission de l’information douloureuse entre les fibres sensorielles périphériques et les structures supraspinales.
La mise en jeu des contrôles segmentaires spinaux a été modélisée par MELZACK et WALL en 1965 dans leur « théorie du portillon » (« gate control theory » en anglais).
Ce modèle repose sur l’équilibre d’une balance entre deux types d’activités d’origine segmentaire périphérique exercées sur les neurones nociceptifs non spécifiques :
– Les unes sont activatrices véhiculées par les fibres nociceptives de petit diamètre ;
– Les autres sont inhibitrices véhiculées par les fibres non nociceptives de la sensibilité tactile légère de gros diamètre.
La douleur n’est ressentie que lorsque la balance penche en faveur des activités excitatrices.
L’activation des fibres non nociceptives fermerait le portillon et bloquerait la transmission de l’information nociceptive vers les structures de la douleur dans l’encéphale (idée associée à une analgésie) ; l’activation des fibres nociceptives ouvrirait le portillon et favoriserait la transmission de l’information nociceptive vers les structures de la douleur dans l’encéphale (idée associée à la sensation de douleur).
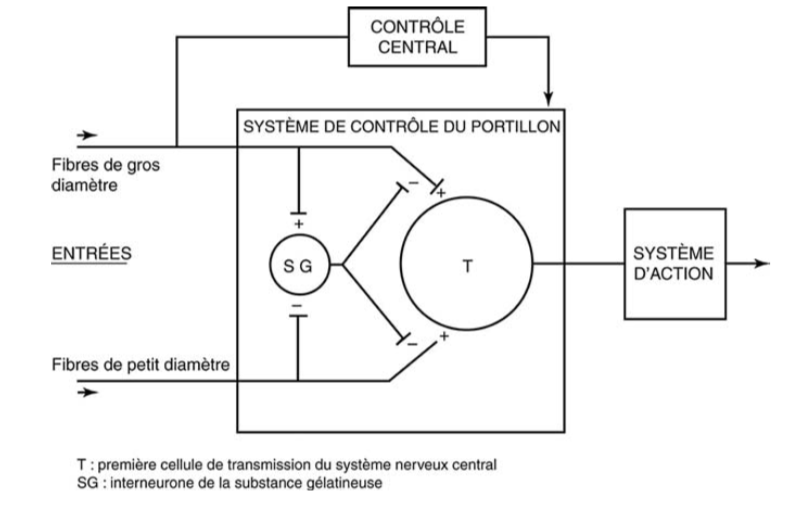
Schéma de la « théorie du portillon » (« gate control theory ») tel que proposé par Melzack et Wall [4] en 1965.
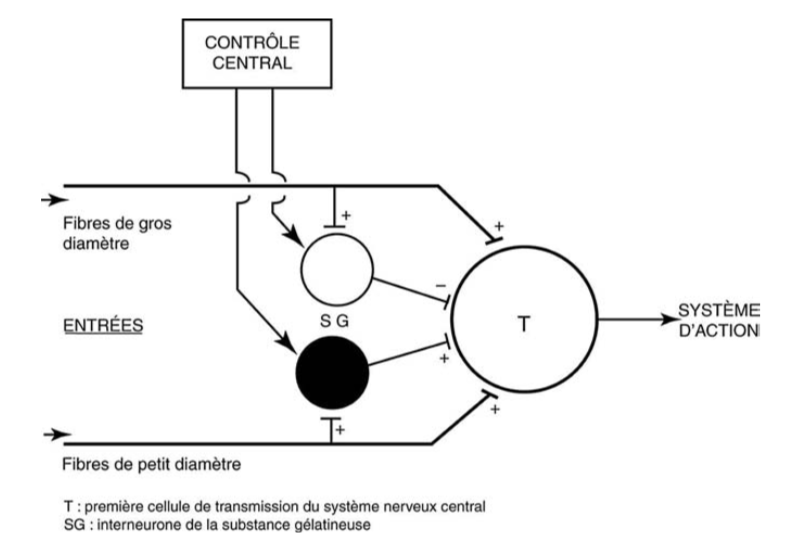
Schéma révisé de la « théorie du portillon ».Deux familles d’ interneurones (l’ une inhibitrice et l’ autre activatrice), et non plus une comme dans le schéma initial, sont mises en jeu dans le système de contrôle.
Dans le modèle du «gate control » le mécanisme de régulation spinal est lui-même soumis à des contrôles descendants d’origine supraspinale (voir ci-dessous).
Ce modèle est bien loin de rendre compte de toutes les données expérimentales mais il a été à l’origine de l’utilisation thérapeutique d’une technique de neurostimulation électrique périphérique transcutanée (« Transcutaneous Electrical Nerve Stimulation », TENS) :
Une stimulation électrique antalgique de faible intensité et de haute fréquence est délivrée par l’intermédiaire d’électrodes de contact cutanées disposées sur le territoire douloureux.
Les effets analgésiques de la TENS, utilisée principalement dans les cas de douleurs chroniques neuropathiques consécutives à des lésions des nerfs sensoriels, sont manifestes après plusieurs minutes de stimulation et peuvent parfois se prolonger au delà de l’arrêt de la stimulation, ce qui n’est pas en accord total avec le modèle !
L’intérêt clinique de cette technique est discuté car son efficacité ne concerne qu’une partie des patients douloureux chroniques et l’on observe une dissipation des ses effets au cours du temps par un effet de tolérance.
Les contrôles inhibiteurs descendants (CID)
Les contrôles inhibiteurs descendants (CID) sont issus de structures du tronc cérébral dont l’origine est la substance grise périaqueducale, SGPA, mésencéphalique :
La stimulation de ces neurones et de ceux des structures en aval sérotoninergiques (neurones des noyaux du Raphé Magnus, des noyaux paragigantocellulaire et gigantocellulaire de la medulla rostro-ventrale, RVM) est à l’origine d’effets analgésiques résultant de la mise en jeu de voies descendantes exerçant un contrôle inhibiteur sur les neurones nociceptifs non spécifiques de la CDME, bloquant la transmission des messages nociceptifs.
Les axones descendants de ces neurones se projettent à tous les différents segments de la moelle épinière (du segment cervical au segment sacré) dans la corne dorsale de la moelle.
On y associe les systèmes de contrôles inhibiteurs descendants noradrénergiques (neurones du locus coeruleus et du locus subcoeruleus), qui fonctionnent sur le même modèle.
D’où l’utilisation de molécules ayant une action centrale à la fois sérotoninergique et noradrénergique, comme certains antidépresseurs, pour obtenir des effets analgésiques dans la douleur chronique neuropathique, qui renforcent ces CID.
Mais des contrôles facilitateurs descendants pro-algiques, également issus du tronc cérébral, ont été décrits exacerbant les conséquences d’une stimulation nociceptive au niveau de la moelle épinière.
Une stimulation de la RVM à des intensités élevées déclenche des effets analgésiques (cf. ci-dessus), mais à des intensités plus faibles dans la même région, déclenche des effets facilitateurs pro-algiques, avec une discrimination anatomique entre des sites inhibiteurs antalgiques et d’autres facilitateurs pro-algiques.
L’équilibre entre ces deux systèmes descendants concurrents déterminerait in fine le degré global d’excitabilité du réseau de neurones dans la CDME, degré qui à son tour modulerait la transmission de l’information douloureuse vers les structures nerveuses centrales de l’encéphale.
Lorsque les deux systèmes sont à l’équilibre, l’organisme est dans un état d’homéostasie physiologique qui se traduit par un état de bien être.
Les contrôles inhibiteurs diffus induits par une stimulation nociceptive (CIDN)
Les contrôles inhibiteurs diffus induits par une stimulation nociceptive (CIDN) constituent le support neurophysiologique de la contre-irritation, qui résulte des effets réciproques entre deux stimulations nociceptives d’intensités différentes, processus par lequel une douleur peut masquer une autre douleur.
Ces CIDN peuvent être déclenchés par une stimulation depuis n’importe quel territoire corporel (stimulation hétérotopique) distinct des territoires activés par la douleur, y compris à partir d’un territoire viscéral, à la condition que cette stimulation hétérotopique soit douloureuse.
Cette vieille pratique clinique était utilisée il y a encore peu de temps (ex. : cataplasmes sinapisés) et, relevant du même principe de contre-irritation, des techniques d’hyperstimulation (ex. : électro-acupuncture) sont aussi utilisées pour obtenir des analgésies, d’autant plus efficaces qu’elles sont d’une intensité à la limite de la tolérance de la douleur du sujet.
Interprétation hypothétique de l’activité globale des neurones spinaux et trigéminaux liés à la nociception.
A : l’ensemble de cette activité pourrait ne pas être négligeable chez les sujets en action, donnant lieu à une « activité somesthésique de base » transmise vers les centres nerveux supérieurs.
B : un stimulus nociceptif active les neurones nociceptifs spécifiques et non spécifiques.
C : ceux-ci vont envoyer un message excitateur au cerveau.
D : ce message déclenche secondairement les CIDN.
E : il en résulte une réduction importante de l’activité des neurones convergents non directement concernés par le stimulus initial.
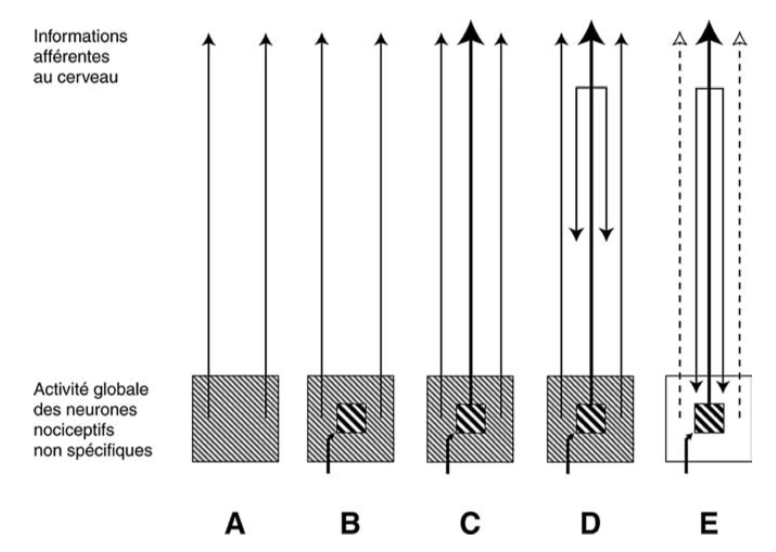
Interprétation hypothétique de l’activité globale des neurones spinaux et trigéminaux liés à la nociception.
D’après Calvino B et Grilo RM. Le contrôle central de la douleur. Revue du rhumatisme 73 (2006) 10-18
Les contrôles issus de diverses structures cérébrales
Plus récemment, des contrôles issus de diverses structures cérébrales ont été décrits, en particulier de différentes aires du cortex et du système limbique.
Lorsqu’elle se prolonge dans le temps, la douleur devient chronique et est à l’origine de mécanismes de sensibilisation, tant périphérique que centrale, qui vont modifier profondément l’activité des systèmes physiologiques de la douleur décrits ci-dessus.
Neuroplasticité : Sensibilisation périphérique et centrale
Ces modifications résultent d’une propriété fondamentale du système nerveux : la neuroplasticité, c’est à dire sa capacité à modifier son activité synaptique et sa morphologie cellulaire, par exemple par bourgeonnement des ses terminaisons axonales centrales.
La sensibilisation périphérique résulte du prolongement dans le temps des processus lésionnels périphériques et des modifications intervenant dans les terminaisons périphériques des nerfs sensoriels du fait de la présence de la soupe inflammatoire :
Les molécules de la soupe inflammatoire sont à l’origine de l’abaissement du seuil de réponse des nocicepteurs et de la potentialisation de l’activité électrique des fibres qui va être exacerbée.
La sensibilisation centrale résulte de cette double propriété des fibres sensorielles et s’exprime par une activation considérable des neurones nociceptifs post-synaptiques de la CDME.
De très nombreux facteurs interviennent dans la mise en place de cette sensibilisation centrale, en particulier la transmission glutamatergique qui y joue un rôle important par l’intermédiaire de l’un des récepteurs du glutamate (un neurotransmetteur activateur abondamment libéré par les fibres nociceptives périphériques chroniquement activées dans les synapses du premier relai médullaire dans la CDME par les stimulations douloureuses chroniques), le récepteur NMDA.
Le récepteur NMDA est un récepteur-canal perméable aux cations, (principalement le calcium) exprimé par les neurones nociceptifs de la CDME, dont la kétamine est un antagoniste (d’où son utilisation comme antalgique).
Le BDNF (Brain Derived Neurotrophic Factor) un neuropeptide synthétisé par les fibres afférentes sensorielles nociceptives, stocké dans leurs terminaisons centrales dans la CDME et libéré par les stimulations chroniques de ces fibres, jouerait un rôle de neuromodulateur dans la douleur chronique. Il pourrait réguler l’excitabilité des neurones nociceptifs post-synaptiques de la CDME.
En conclusion, la douleur chronique résulte de l’expérience subjective d’une sensation émotive déplaisante.
Chacun d’entre nous est unique face à sa douleur :
La douleur chronique peut être considérée comme un langage individuel, une des caractéristiques les plus signifiantes de notre personnalité.
L’Association Internationale d’Etude de la Douleur (International Association for the Study of Pain – IASP) définit la douleur comme « une expérience sensorielle et émotionnelle désagréable associée à une atteinte tissulaire réelle ou potentielle ou décrites en termes d’un tel dommage ».
Cette définition présente l’immense avantage de parler d’une expérience émotionnelle et non uniquement d’une sensation mesurable.
Le contrôle de la douleur est un problème qui va bien au delà de la mise en jeu de réseaux de neurones, segmentaires et/ou descendants d’origine supra-spinale, car il prend aussi en compte les facteurs psychologiques et la personnalité de chacun dans sa globalité.
Mais toute la complexité du problème vient du fait que la douleur chronique, dans le passage de la physiologie de la nociception à la pathologie de la douleur chronique, remet en question le fonctionnement des réseaux neuronaux et synaptiques de la nociception du fait de la neuroplasticité du système nerveux central.
On ne peut réduire la douleur au seul aspect anatomo-clinique de la nociception, car fondamentalement le processus thérapeutique du traitement de la douleur chronique a pour but de prendre en charge non plus la douleur mais l’homme douloureux : « guérir de sa douleur plutôt que guérir sa douleur. »



